Noot van de redactie: Dit artikel is al een wat ouder artikel (2002). De argumentatie in dit artikel is nog steeds bruikbaar en waardevol. Het zou goed zijn wanneer een sedimentoloog, een geochemicus en een bacterioloog samen zouden werken aan de verdere uitwerking en aanvulling van dit argument, daarbij gebruikmakend van nieuwe gegevens uit het veld.
Samenvatting
In veel sedimenten worden levende bacteriën gevonden, evenals bacteriën in slaaptoestand, bekend als sporen. De bacteriën kunnen afgezet zijn tijdens de sedimentatie (autochtone oorsprong), maar dit is vaak moeilijk te bewijzen, omdat de mogelijkheid van later binnendringen in het sediment (allochtone oorsprong) of besmetting van de monsters uitgesloten moet worden. De gepubliceerde ouderdom van de betreffende sedimenten lopen op tot honderden miljoenen jaren.
Levensvatbare sporen, met bewezen autochtone oorsprong, zijn recent verkregen uit waterdruppeltjes die ingesloten zijn in de Salado-zoutlaag (evaporiet) uit het late Perm, in New Mexico (USA). Dit evaporiet wordt gedateerd op 250 miljoen jaar (Ma). Deze ouderdom is echter niet in overeenstemming met bekende vervalsnelheden van bacterieel leven (dat wil zeggen: de snelheid waarmee het aantal levende of levensvatbare individuen afneemt gedurende de tijd). Integendeel, deze vervalsnelheden geven aan dat de ouderdom van het Salado-evaporiet uit het late Perm minder is dan 35.000 jaar.
Schattingen van ouderdom op basis van bacteriële vervalsnelheden zijn kwalitatief, of hooguit semi-kwantitatief. Toch zou de schatting van de ouderdom van het Salado-evaporiet verbeterd kunnen worden door een schatting te maken van de gemiddelde temperatuur van het Salado-evaporiet gedurende de tijd, in combinatie met het vaststellen van de vervalsnelheid van de bacterie Bacillus marismortui, onder condities die representatief zijn voor waterinsluitingen.

Hedendaagse bacteriën
Bacteriën worden vrijwel overal op aarde gevonden en vele soorten overleven, of floreren zelfs, onder extreme condities, zoals in water bij verhoogde druk tot boven 110 ˚C, in ijs beneden -10 ˚C, in zuren bij pH waarden tot 2 en in basen bij pH waarden tot 13, en in milieus zonder zuurstof. Zij kunnen zich met eigenlijk alles voeden, inclusief anorganische voedingsstoffen die vrijkomen rond middenoceanische hete bronnen, maar ook in afwezigheid van voeding kunnen zij lange periodes overleven in de vorm van sporen.
De aanwezigheid van levende bacteriën in sedimenten wordt al bijna een eeuw onderzocht. Allochtone of autochtone oorsprong, de beschikbaarheid van voedingsstoffen gedurende lange tijdsperiodes, en de mogelijkheid van besmetting van monsters gedurende verzameling en behandeling zijn nog steeds onderwerp van discussie. Gedurende de jaren ’70 werden in de sedimenten van een meer levensvatbare sporen van een thermofiele (warmteminnende) bacterie ontdekt met radiokoolstofouderdom van 7.000 jaar en ouder. De mogelijkheid dat deze sedimenten op een later tijdstip besmet waren met deze bacterie kon uitgesloten worden, omdat de sedimenten nooit boven 35 ˚C zijn geweest, wat de minimale temperatuur is voor het doen ontwaken van de sporen van deze specifieke soort thermofiele bacterie. Een ander bekend voorbeeld zijn de bacteriën verkregen uit de ingewanden van een bevroren Siberische mastodont, gedateerd op 11.000 jaar. Deze en andere studies geven aan dat, in afwezigheid van metabolische activiteit, de leeftijd van autochtone bacteri?n aanwezig in recente sedimenten kan oplopen tot ongeveer 10.000 radiokoolstofjaren.
Oeroude bacteriën
Een punt van discussie is de leeftijd van bacteriën die zijn ingesloten in pre-Pleistocene Fanerozoïsche sedimenten. Meerdere malen is de aanwezigheid van bacteriën diep onder de oppervlakte van de aarde gerapporteerd, in verschillende typen sediment, inclusief koolsteen en oliehoudende lagen en in zoutlagen met gerapporteerde ouderdom tot 650 miljoen jaar. De bewijsvoering voor de autochtone oorsprong is niet algemeen geaccepteerd, omdat het moeilijk is om allochtone oorsprong of besmetting gedurende verzameling en behandeling uit te sluiten.123 De aanwezigheid van bacteriën in afgelegen oliebronnen is bijvoorbeeld vastgesteld, waarbij de oorsprong van deze bacteriën niet werd opgelost. Zij zouden ingesloten kunnen zijn tijdens de vorming van de olie, ze kunnen meegevoerd zijn met ondergrondse waterstromen of ze zouden zich zelfstandig verplaatst kunnen hebben gedurende tientallen tot honderden miljoenen jaren.4
Recent werden levensvatbare sporen van Bacillus sphaericus geïsoleerd uit een in amber ingesloten uitgestorven bijensoort met een gerapporteerde ouderdom van 25?30 miljoen jaar.5 De zorgvuldige selectie van het monster en de grondige voorzorgsmaatregelen die werden genomen tegen besmetting heeft deze vondst overtuigend gemaakt.
Nog recenter werden bacteriën verkregen uit autochtone waterinsluitingen in zoutmonsters van de Permische Salado-evaporiet (New Mexico, USA) met een gerapporteerde ouderdom van 250 miljoen jaar.6 Deze oude bacteri?n hebben een opmerkelijke overeenkomst met Bacillus marismortui (vernoemd naar de Dode Zee) en Virgobacillus pantothenticus, gebaseerd op de vergelijking van 16S-ribosomaal RNA. Het verschil met V. pantothenticus is klein, met een overeenkomst van 97.1% (alhoewel Vreeland 97.5% overeenkomst vermeldt). 45 van de 1650 basenparen zijn verschillend, wat overeenkomt met jaarlijks 1×10^-10 substituties per basenparen. Deze vervangingssnelheid is laag, maar mogelijk aanvaardbaar.7 De 99% overeenkomst met B. marismortui is echter nog opmerkelijker.
Als deze oude bacteriën inderdaad familie zijn van B. marismortui, wat nog vastgesteld moet worden, hoe kan dit verschil dan zo klein zijn? De vrij in de natuur levende B. marismortui zou al gedurende 250 miljoen jaar mutaties moeten ondergaan en we mogen een veel groter verschil met de geïsoleerde soortgenoten verwachten89, alhoewel een efficiënt DNA reparatiemechanisme, actief gedurende het ontwaken, de overeenkomst misschien gedeeltelijk zou kunnen verklaren.10
De maximale leeftijd van bacteriën is in dit verband bepalend en zal nu in meer detail uitgewerkt worden.11
Het overleven van bacteriën
Als er voldoende voeding beschikbaar is kunnen bacteriën in principe oneindig lang blijven leven. Zij vermenigvuldigen zich aseksueel door deling (een cel verdeelt zich in twee cellen) en zowel de moeder- als de dochtercellen (alhoewel die niet van elkaar te onderscheiden zijn) hebben zich vernieuwd, waardoor de populatie haar levenskracht behoud, mits er optimale voeding aanwezig blijft en het niveau van uitgescheiden toxische verbindingen laag is. In afwezigheid van voeding is de levensduur van bacteri?n beperkt, maar er kan geen bepaalde maximale levensverwachting vastgesteld worden. Als de hoeveelheid voeding afneemt of als het niveau van de uitgescheiden toxische verbindingen toeneemt in de loop van de tijd, neemt het aantal levende individuen geleidelijk af. De snelheid waarmee het bacteriële leven vervalt wordt gewoonlijk uitgedrukt als de decimale reductietijd, D(10), de tijdsduur bij een bepaalde temperatuur waarin het aantal levende bacteriën in een populatie tot 10% is teruggelopen. Deze snelheid is vergelijkbaar met andere exponentieel afnemende processen, zoals eerste-orde chemische reacties of radioactief verval.
Bacteriën kunnen gedroogd en onder vacuüm ongeveer een jaar bewaard worden, of gedurende enkele jaren in vloeibare stikstof (-196 ˚C).12 Zij kunnen gedurende veel langere tijden overleven als ze enige metabolische activiteit kunnen onderhouden. Bacteriële culturen zijn aangetroffen in Siberische permafrost bij een gemiddelde temperatuur rond -10 ˚C. De duur van de permafrost wordt geschat tussen duizend en een miljoen jaar. Er werd verondersteld dat de bacteriën actief gebleven moeten zijn ondanks de lage temperatuur, omdat zij anders niet zo lang konden overleven.13 De recente ontdekking van bacteriën in ijs en sneeuw op Antarctica biedt ondersteuning voor lage-temperatuuractiviteit van bacteriën. Deze gevonden bacteri?n hebben inderdaad metabolische activiteit bij -12 ˚C tot -17 ˚C.14
Het is interessant om op te merken dat de Antarctische onderzoekers leden van Deinococcus ontdekten, een geslacht van bacteriën dat bekend is in de voedingsmiddelenindustrie vanwege haar extreme vermogen om ioniserende straling te weerstaan, inclusief röntgenstraling en radioactiviteit.15 Dit is opmerkelijk, omdat zulke hoge stralingsniveaus in een natuurlijke omgeving niet voorkomen en het onduidelijk was welk voordeel dit vermogen zou kunnen bieden. Het antwoord zou kunnen zijn dat Deinococcus-soorten blootgesteld kunnen staan aan intense UV-straling, waardoor eveneens ionisatie optreedt, bijvoorbeeld in de sneeuwvelden van Antarctica.16 Dit voorbeeld laat opnieuw zien dat sommige bacteriën de opmerkelijke eigenschap hebben om onder extreme condities te overleven.
De meeste bacteriën hebben moeite om te overleven als zij worden onderworpen aan een hoge temperatuur, straling (inclusief UV straling), oxidatoren (inclusief zuurstof), een aantal anorganische componenten, mechanische krachten en natte omstandigheden.17 Echter in een waterinsluiting in een evaporiet kunnen veel van deze condities voor bacteriën gunstig zijn, zelfs als er water aanwezig is. Hoewel zout desastreus is voor de meeste bacteriën, bestaan er uitzonderingen zoals B. marismortui, die kan overleven als sporen of zelfs floreren onder zoute condities bij concentraties beneden 25% w/v.
Samenvattend: sommige bacteriën kunnen extreme condities weerstaan mits enige metabolische activiteit kan plaatsvinden. In afwezigheid van voedingsstoffen kan de levensduur van bacteriën jaren of tientallen jaren bedragen, maar het overleven gedurende duizenden jaren in afwezigheid van metabolische activiteit is iets te veel gevraagd. Kunnen bacteriën 250 Ma overleven als zij opgesloten zijn in een waterinsluiting in zout?
De ‘bacteriële ouderdom’ van de Salado-evaporiet.
Bacillus marismortui vormt sporen als de zoutconcentratie hoger wordt dan 25% w/v. De verwachting is dat de sporen niet ontwaken in een verzadigde zoutoplossing, ook niet in aanwezigheid van voeding. Het aantal levensvatbare sporen in de Salado-afzetting is in de loop van de tijd gestaag afgenomen, van mogelijk oorspronkelijk 100 miljoen bacteri?n per ml (ref. 2) tot slechts enkele sporen in 3 van de 66 onderzochte insluitingen, met een tot op heden totaal volume (van de 66 insluitingen) van 0,365 ml.18 Als een minimum van drie levensvatbare sporen in 0,365 ml wordt verondersteld betekent dit dat er zeven periodes van decimering zijn gepasseerd. Met een D(10) van 50 jaar betekent dit een ‘bacteri?le ouderdom’ van 350 jaar voor de Salado-evaporiet. Aanvullend ondersteunend bewijs voor de jonge leeftijd van deze evaporiet is de hierboven genoemde overeenkomst tussen 16S ribosomaal RNA van de gevonden oerbacterie en B. marismortui of V. pantothenticus.
Het verschil met de gepubliceerde ouderdom van 250 Ma voor de Salado-evaporiet is bijna zes ordegroottes! Een ongeloofwaardige D(10) van 35 Ma in plaats van 50 jaar is nodig om het totaal van 250 Ma te bereiken. Omgekeerd, uitgaande van een ouderdom van de evaporiet van 250 Ma en een decimeringstijd van 50 jaar betekent dat de oorspronkelijk bacteriën vijf miljoen decimeringsperiodes hebben doorgemaakt. Het is onmogelijk om ook maar een enkel individu terug te vinden na vijf miljoen decimeringen.
Het verschil met de gepubliceerde ouderdom van 250 Ma voor de Salado-evaporiet is bijna zes ordegroottes! Een ongeloofwaardige D(10) van 35 Ma in plaats van 50 jaar is nodig om het totaal van 250 Ma te bereiken. Omgekeerd, uitgaande van een ouderdom van de evaporiet van 250 Ma en een decimeringstijd van 50 jaar betekent dat de oorspronkelijk bacteriën vijf miljoen decimeringsperiodes hebben doorgemaakt. Het is onmogelijk om ook maar een enkel individu terug te vinden na vijf miljoen decimeringen.
Een ouderdom van 350 jaar voor de Salado-evaporiet valt binnen de historische tijd en is natuurlijk niet realistisch. Dit komt omdat er rekening gehouden moet worden met een groot aantal onzekerheden.
Het kleine aantal levensvatbare sporen (minimaal drie) heeft statistisch gezien nauwelijks enige waarde, en het resultaat is een brede reeks van mogelijke ouderdommen, waarbij hogere ouderdom meer waarschijnlijk is dan lagere ouderdom.
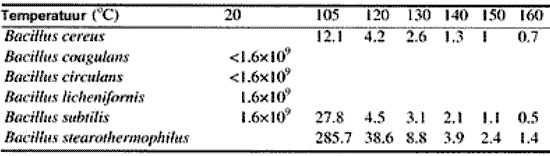
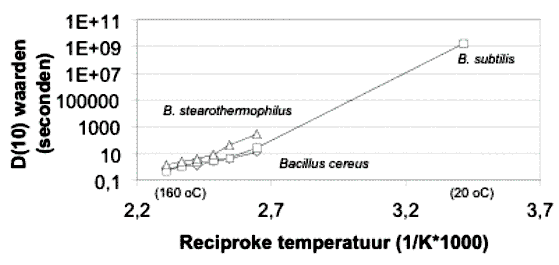
De D(10)-waarde van B. marismortui onder condities die representatief zijn voor waterinsluitingen is onbekend en moet apart vastgesteld worden. De decimeringstijden variëren met ongeveer een orde van grootte voor verschillende soorten (Tabel 1). Een verandering van temperatuur met 25 ˚C heeft een effect van twee ordegroottes op de decimeringstijd (Figuur 1).
Een gemiddelde temperatuur rond of beneden 0 ˚C gedurende de levensduur van de evaporiet is buitengewoon onwaarschijnlijk. Een gemiddelde temperatuur rond of boven 20 ˚C kan mogelijk zijn, met mogelijk kortere D(10) tijden en dus een lagere ouderdom voor de evaporiet.
De berekende ‘bacteriële ouderdom’ van 350 jaar is daarom erg onzeker en zou makkelijk met twee of misschien drie ordes van grootte kunnen variëren, met een hogere waarschijnlijkheid voor hogere dan voor lagere ouderdom. Dit resulteert in een geschatte bandbreedte van 200 tot 35.000 jaar, corresponderend met 4 log cycli van 50 jaar bij 20 ˚C of 7 log cycli en een D(10) van 5.000 jaar bij 0 ˚C, respectievelijk. Het kost moeite om zich condities voor te stellen die tot een ouderdom van 350.000 jaar zouden leiden, maar zelfs deze schatting verschilt meer dan twee ordegroottes met de 250 Ma die gepubliceerd is voor de Salado-evaporiet. Dit verschil zou nog groter kunnen worden als beweringen voor 650 Ma oude bacteriën in evaporieten bevestigd zouden worden.
Het is verrassend dat de aanwezigheid van levensvatbare autochtone bacteriële sporen in de Salado-evaporiet niet heeft geleid tot het ter discussie stellen van haar geologische leeftijd. Parkes geeft de voorkeur aan de veronderstelling dat bacteriën vrijwel onsterfelijk zijn, boven het in twijfel trekken van de geologische tijdschaal. Zelfs het overleven van bacteriën in meteorieten en de oorsprong van leven op aarde via interplanetair transport van bacteriële sporen (‘pansperma’) worden overwogen. Meer recent ontkende Lindahl de hoge ouderdom van de verkregen bacteriën op basis van de onmogelijkheid dat DNA langer dan 100.000 jaar zou kunnen overleven, met als conclusie dat deze bacteri?n de hedendaagse algemeen voorkomende bacteriën zijn, dus van allochtone oorsprong.19
Conclusies
De aanwezigheid van levensvatbare sporen in water insluitingen in de Permische Salado-evaporiet is niet in overeenstemming met de gepubliceerde ouderdom van 250 Ma.
De ‘bacteriële ouderdom’ van de Permische Salado-evaporiet is semi-kwantitatief geschat en ligt waarschijnlijk tussen 350 en 35.000 jaar, onder de aanname dat gepubliceerde bacteriële vervalsnelheden representatief zijn voor de omstandigheden in waterinsluitingen in een evaporiet.
De schatting op basis van de ‘bacteriële ouderdom’-methode kan verder worden verbeterd als de gemiddelde temperatuur van de evaporiet geschat zou kunnen worden en als de decimeringstijden van B. marismortui over een brede temperatuurrange gemeten zou worden.
Deze gastbijdrage is vertaald door leden van <em)Mediagroep in Genesis en met toestemming overgenomen van de website Schepping-of-Evolutie. Het originele artikel is hier te vinden. Het artikel is eerder verschenen in Technical Journal (tegenwoordig Journal of Creation) van Creation Ministries International (CMI). De bronvermelding luidt: Heide, E.A. van der, 2002, Bacterial Life in Ancient Salt, Technical Journal 16 (2): 110-113 (artikel).
Referenties en aantekeningen
- Postgate, J., The outer reaches of life, University Press, Cambridge, 1994.
- Parkes, R.J. (2000), A case of bacterial immortality?, Nature 407 (6806): 844-845, 19 October 2000.
- Roth, A.A., Origins, Review and Herald Publishing Association, Hagerstown, p. 164, 1998. Dit boek is door drs. Pieter Bouma in het Nederlands vertaald en verschenen bij Uitgeverij Groen.
- Magot, M., Similar bacteria in remote oil fields, Nature 379 (6567): 681, 22 February 1996.
- Cano, R.J. and Borucki, M., Revival and identification of bacterial spores in 25-to 40-million-year old Dominican amber, Science 268 (5213): 1060-1064, 19 May 1995.
- Vreeland, R.H., Rosenzweig, W.D. and Powers, D.W., Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal, Nature 407 (6806): 897-900, 19 October 2000.
- Hartl, D.L. and Clark, A.G., Principles of Population Genetics, Sinauer, Sunderland, p. 107, 1989.
- Parkes, R.J. (2000), A case of bacterial immortality?, Nature 407 (6806): 844-845, 19 October 2000.
- Vreeland, R.H., Rosenzweig, W.D. and Powers, D.W., Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal, Nature 407 (6806): 897-900, 19 October 2000.
- Nicholson, W.L., Munakata, N., Horneck, G., Melosh, H.J. and Setlow, P., Resistance of Bacillus endospores to extreme terrestrial and extraterrestrial environments, Microbiology and Molecular Biology Reviews 64(3):548?572, September 2000.
- Van der Heide, E., Leven in aardlagen, Bijbel en Wetenschap 225: 5-8, 2000.
- Schlegel, H.G., General Microbiology, 7th ed., Cambridge, 1993.
- Shi, T., Reever, R.H., Gilchinsky, D.A. and Friedman, E.I., Characterisation of viable bacteria from Siberian Permafrost by 16S rDNA sequencing, Microbial Ecology 33 (3): 169-179, 1997.
- Carpenter, E.J., Lin, S. and Capone, D.G., Bacterial Activity in South Pole Snow, Applied and Environmental Microbiology 66 (10): 4514-4517, October 2000.
- Makarova, K.S., Aravind, L., Wolf, Y.I., Tatusov, R.L., Minton, K.W., Koonin, E.V. and Daly, M.J., Genome of the extremely radiation-resistant bacterium Deinococcus radiodurans viewed from the perspective of comparative genomics, Microbiology and Molecular Biology Reviews 65 (1): 44-79, March 2001.
- Carpenter, E.J., Lin, S. and Capone, D.G., Bacterial Activity in South Pole Snow, Applied and Environmental Microbiology 66 (10): 4514-4517, October 2000.
- Schlegel, H.G., General Microbiology, 7th ed., Cambridge, 1993.
- Vreeland, R.H., Rosenzweig, W.D. and Powers, D.W., Isolation of a 250 million-year-old halotolerant bacterium from a primary salt crystal, Nature 407 (6806): 897-900, 19 October 2000.
- Lindahl, T., Unlocking nature’s ancient secrets: review of: ‘The Molecule Hunt: Archaeology and the Search for Ancient DNA’ by Martin Jones, Nature 413 (6854): 358-359, 27 September 2001.